NCERT Book Class 12 Biology Chapter 2 Human Reproduction is here. You can read and download Class 12 Biology Chapter 2 PDF from this page of aglasem.com. Human Reproduction is one of the many lessons in NCERT Book Class 12 Biology in the new, updated version of 2023-24. So if you are in 12th standard, and studying Biology textbook (named Biology), then you can read Ch 2 here and afterwards use NCERT Solutions to solve questions answers of Human Reproduction.
NCERT Book Class 12 Biology Chapter 2 Human Reproduction
The complete Chapter 2, which is Human Reproduction, from NCERT Books for Class 12 Biology is as follows.
NCERT Book Class 12 Biology Chapter 2 Human Reproduction PDF Download Link – Click Here To Download The Complete Chapter PDF
NCERT Book Class 12 Biology Full Book PDF Download Link – Click Here To Download The Complete Book PDF
NCERT Book Class 12 Biology Chapter 2 Human Reproduction PDF
The direct link to download class 12 Biology NCERT Book PDF for chapter 2 Human Reproduction is given above. However if you want to read the complete lesson on Human Reproduction then that is also possible here at aglasem. So here is the complete class 12 Biology Ch 2 Human Reproduction.
NCERT Book Class 12 Biology Chapter 2 Human Reproduction View Download
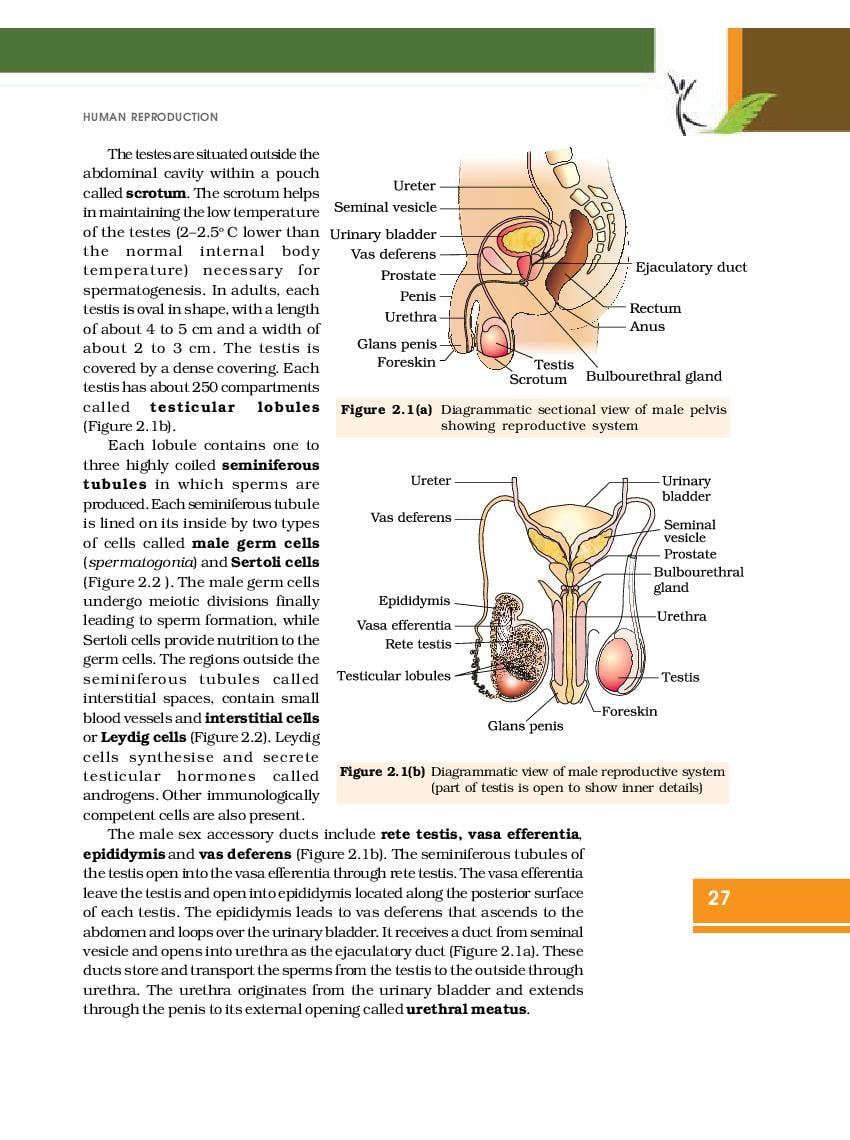
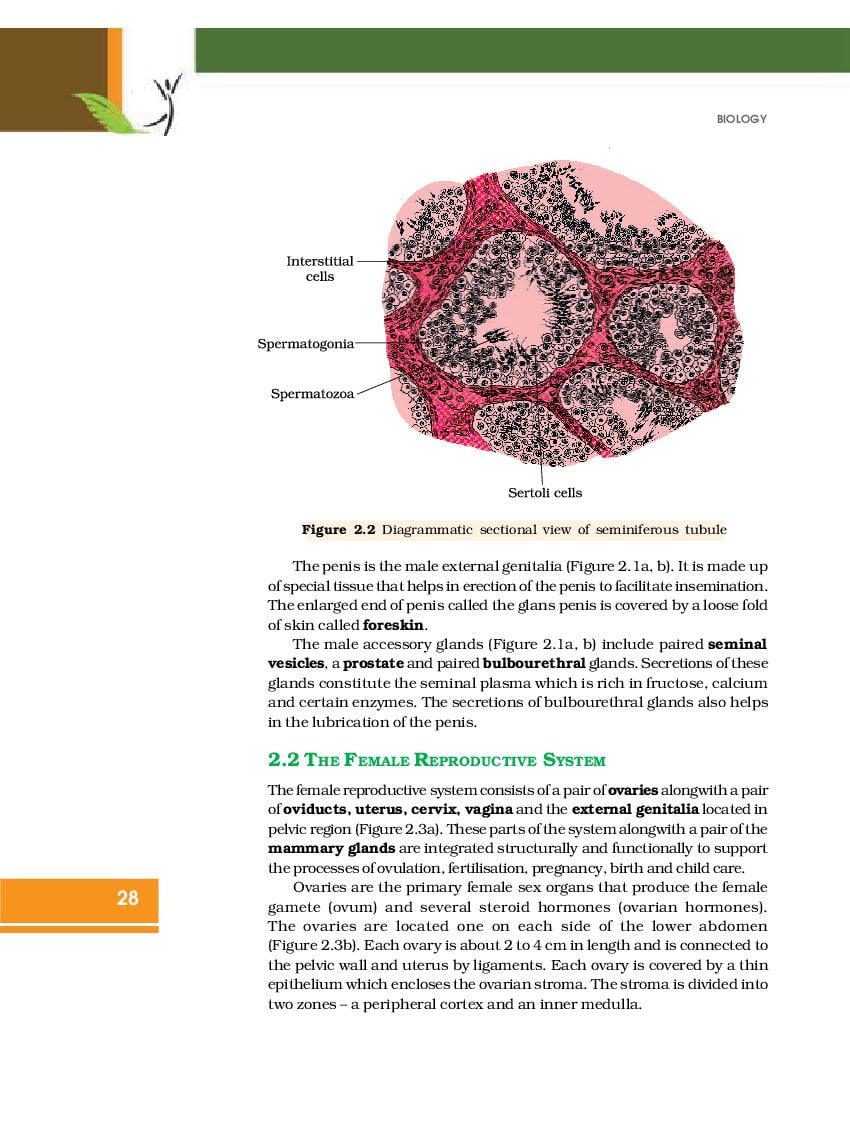
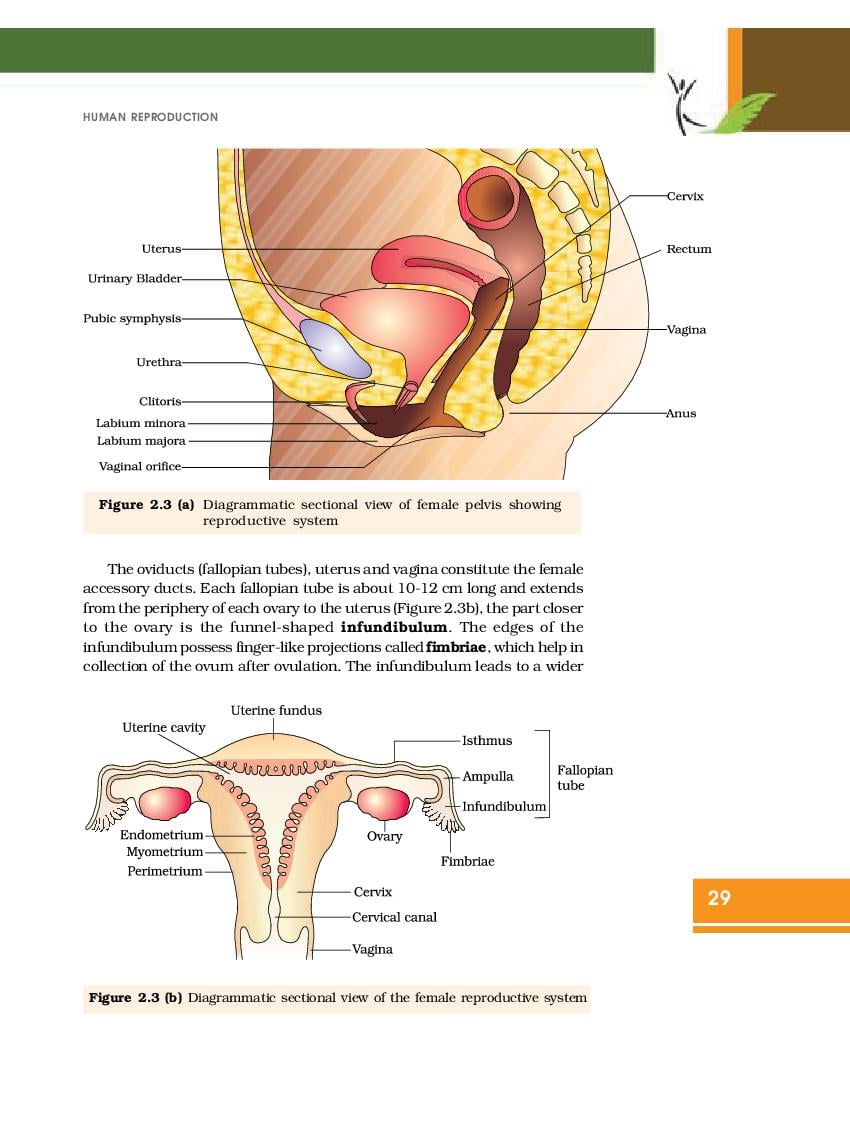
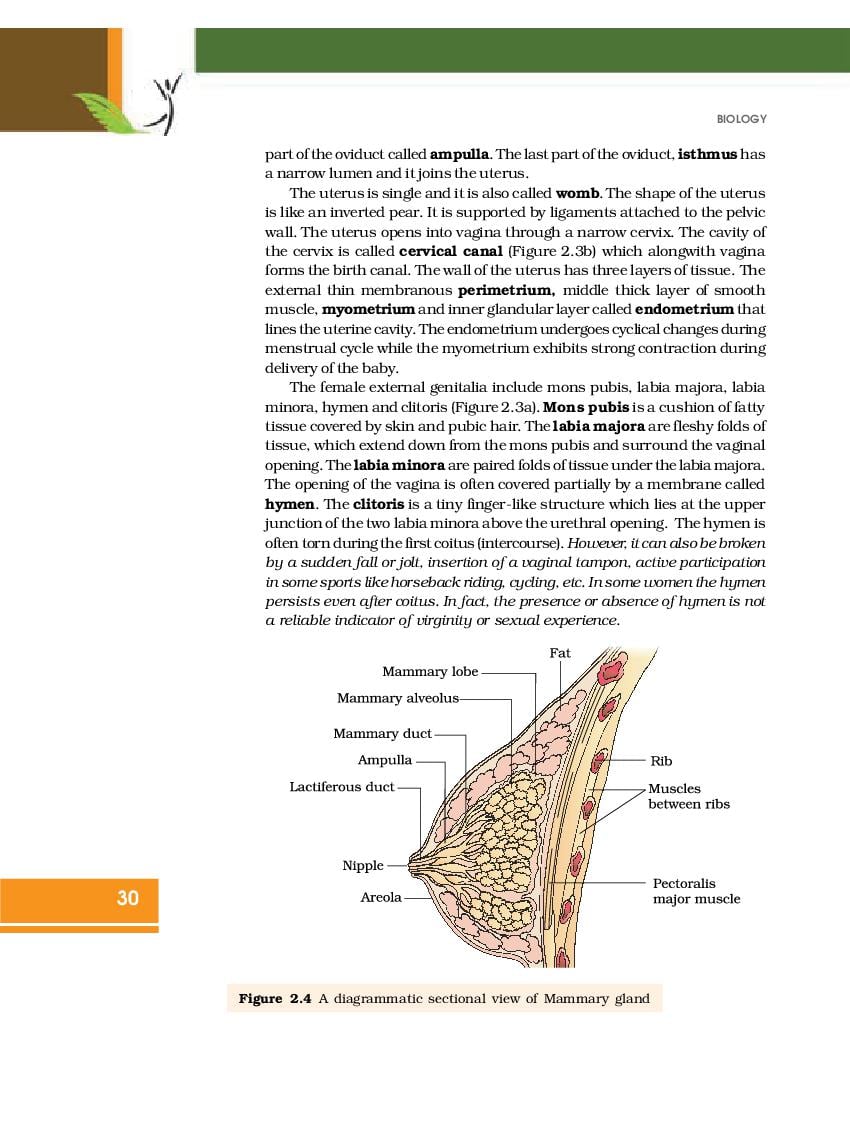








NCERT Book for Class 12 Biology
Besides the chapter on Human Reproduction, you can read or download the NCERT Class 12 Biology PDF full book from aglasem. Here is the complete book:
- Chapter 1 Reproduction in Organisms
- Chapter 2 Sexual Reproduction in Flowering Plants
- Chapter 3 Human Reproduction
- Chapter 4 Reproductive Health
- Chapter 5 Principles of Inheritance and Variation
- Chapter 6 Molecular Basis of Inheritance
- Chapter 7 Evolution
- Chapter 8 Human Health and Disease
- Chapter 9 Strategies for Enhancement in Food Production
- Chapter 10 Microbes in Human Welfare
- Chapter 11 Biotechnology: Principles and Processes
- Chapter 12 Biotechnology and its Applications
- Chapter 13 Organisms and Populations
- Chapter 14 Ecosystem
- Chapter 15 Biodiversity and Conservation
- Chapter 16 Environmental Issues
NCERT Books for Class 12
Similarly all the subject-wise class 12 books at aglasem.com are as follows.
- NCERT Book Class 12 Accountancy
- NCERT Book Class 12 Biology
- NCERT Book Class 12 Business Studies
- NCERT Book Class 12 Chemistry
- NCERT Book Class 12 Economics
- NCERT Book Class 12 English
- NCERT Book Class 12 Geography
- NCERT Book Class 12 Hindi
- NCERT Book Class 12 History
- NCERT Book Class 12 Maths
- NCERT Book Class 12 Physics
- NCERT Book Class 12 Political Science
- NCERT Book Class 12 Psychology
- NCERT Book Class 12 Sociology
NCERT Books
All class-wise books of National Council of Educational Research and Training are as follows.
- NCERT Books for Class 1
- NCERT Books for Class 2
- NCERT Books for Class 3
- NCERT Books for Class 4
- NCERT Books for Class 5
- NCERT Books for Class 6
- NCERT Books for Class 7
- NCERT Books for Class 8
- NCERT Books for Class 9
- NCERT Books for Class 10
- NCERT Books for Class 11
- NCERT Books for Class 12
Class 12 Biology Chapter 2 Human Reproduction NCERT Textbook – An Overview
The highlights of this Human Reproduction chapter PDF are as follows.
| Aspects | Details |
|---|---|
| Class | 12 |
| Subject | Biology |
| Book | Biology |
| Chapter Number | Ch 2 |
| Chapter Name | Human Reproduction |
| Book Portion Here | NCERT Book Class 12 Biology Ch 2 Human Reproduction |
| Download Format | |
| Version | NCERT Book (New, Updated) 2023-24 |
| Complete Book | NCERT Book Class 12 Biology |
| All Class 12 Books | NCERT Books for Class 12 |
| All Textbooks | NCERT Books |
| NCERT Books in Hindi | NCERT Books for Class 12 in Hindi |
| NCERT Solutions | NCERT Solutions for Class 12 |
| More Study Material | Class 12 |
If you have any queries on NCERT Book Class 12 Biology Chapter 2 Human Reproduction, then please ask in comments below. And if you found the Class 12 Biology Chapter 2 Human Reproduction PDF helpful, then do share with your friends on telegram, facebook, whatsapp, twitter, and other social media! :)
To get study material, exam alerts and news, join our Whatsapp Channel.

